
Mécanismes de la contraction musculaire
La théorie qui prévaut actuellement pour expliquer comment les muscles créent des tensions internes (forces) est celle dite "des filaments glissants" développée Huxley (1957) et basée sur le modèle de Hanson et Huxley (1955). Elle stipule que, lors de la contraction musculaire, les filaments d'actine se glissent entre les filaments de myosine. Ce glissement a d'abord été mis en évidence par microscopie électronique (Figure 1).
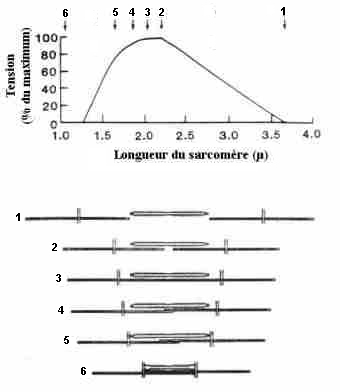
| Figure 1 : Démonstration de la théorie des filaments glissants d'après Gorgon et coll. (1966). |
| A gauche, les images obtenues avec
un microscope électronique. A droite le schéma explicatif
de la théorie des filaments glissants. Chaque chiffre représente
un index de position que l'on peut repérer à la fois sur
les photos (à gauche), sur la courbe tension-longueur (en haut
à droite) et sur le schéma représentatif du processus
qui se déroule lors de la contraction musculaire, c'est-à-dire
le raccourcissement des sarcomères (en bas à droite) lorsque
la résistance extérieure est inférieure à
la tension générée par le sarcomère. La position 2 correspond à la longueur de repos du sarcomère. C'est la longueur optimale pour développer la force la plus élevée car à c'est cette longueur que peuvent se former le plus grand nombre de ponts d'acto-myosine. Or, plus leur nombre est important, plus il y a de possibilités de transformer de l'énergie chimique en énergie mécanique. Pour la position 1, la longueur du sarcomère correspond à 160% de celle mesurée au repos, et les filaments ne se chevauchent plus. Il s'en suit qu'aucun pont d'actine-myosine ne peut se former. Par conséquent, la force que le muscle peut développer est nulle. Par contre, en position 4 (80% de la longueur de repos), le chevauchement des deux myofilaments d'actine entraîne une diminution de la force produite du fait de la réduction de l'accessibilité des sites d'attache aux têtes de myosines. Cet effet s'amplifie pour des longueurs de sarcomère inférieures (75% en position 5 et 45% en position 6). |
Par
la suite, des grossissements plus poussés ont confirmé les premiers
résultats comme le montre la figure 2 lors d'enregistrements réalisés
au cours d'un allongement de l'échantillon de fibres musculaires. C'est
le degré de recouvrement (Figure 2) qui informera sur les possibilités
de production de force (nombres de ponts d'acto-myosine) et surtout de son intensité.
Des relations entre ces grandeurs mécaniques ont pu être mises
en évidence en condition isométrique pour une longueur de fibre
musculaire donnée (relation force-longueur) ou
un angle articulaire donné (relation force-angle)
ou à une vitesse de mouvement donnée (relation
force-vitesse).
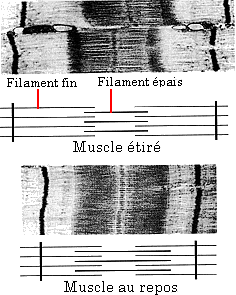
Figure 2 : Mise en évidence
du glissement des filaments
composant le sarcomère par la microscopie électronique
à fort grossissement.
L'animation ci-dessous montre bien que la tension générée par le sarcomère est proportionnelle au taux de recouvrement des myofilaments fins et épais.


Ce glissement est le résultat de la formation de ponts d'union entre les têtes de myosine et certains sites des filaments d'actine déclencher une réaction chimique (hydrolyse de l'ATP) qui libère l'énergie nécessaire l'accrochage des têtes de myosine et, par-là même, d'exercer une traction sur les filaments fins pour les faire glisser entre les filaments épais. Comme le montre l'animation ci-dessus, la force générée est directement proportionnelle au nombre de ponts d'actine-myosine formés. Selon la force extérieure qui s'oppose à la tension ainsi générée, il y aura ou non raccourcissement du muscle. Donc, il se produit dans le muscle une transformation d'énergie chimique en énergie mécanique (Figure 3). Une autre animation est disponible sur le site de l'académie de Rennes.
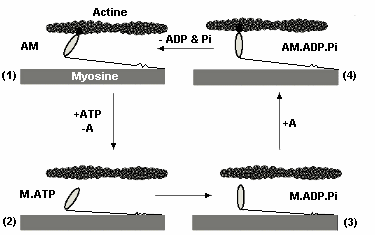
| Figure 3 : Production de la force grâce l'interaction entre l'actine et la myosine. |
1) La myosine (M) est attachée
à l'actine (A) formant ainsi le complexe acto-myosine (AM). |
Bibliographie